 Таким образом, в клетках как гликолиз, так и глюконеогенез являются необратимыми процессами. Необходимо отметить, что гликолиз протекает в цитозоле, а часть реакций глюконеогенеза происходит в митохондриях. Его превращение в пируват и дальнейшее использование последнего являются способом утилизации лактата. Аминокислоты образуются в результате распада мышечных белков и включаются в глюконеогенез при длительном голодании или продолжительной мышечной работе.
Таким образом, в клетках как гликолиз, так и глюконеогенез являются необратимыми процессами. Необходимо отметить, что гликолиз протекает в цитозоле, а часть реакций глюконеогенеза происходит в митохондриях. Его превращение в пируват и дальнейшее использование последнего являются способом утилизации лактата. Аминокислоты образуются в результате распада мышечных белков и включаются в глюконеогенез при длительном голодании или продолжительной мышечной работе.
Некоторые ткани, например мозг, нуждаются в постоянном поступлении глюкозы. Однако запасы гликогена в печени невелики. Глюконеогенез — процесс синтеза глюкозы из веществ неуглеводной природы. Его основной функцией является поддержание уровня глюкозы в крови в период длительного голодания и интенсивных физических нагрузок.
Эти ткани могут обеспечивать синтез 80-100 г глюкозы в сутки. На долю мозга при голодании приходится большая часть потребности организма в глюкозе. Первичные субстраты глюконеогенеза — лактат, аминокислоты и глицерол. Включение этих субстратов в глюконеогенез зависит от физиологического состояния организма. Он образуется при любых состояниях организма в эритроцитах и работающих мышцах. Таким образом, лактат используется в глюконеогенезе постоянно.
Итак, в печени существуют 4 фермента, которые принимают участие только в глюконеогенезе и катализируют обходные реакции необратимых стадий гликолиза
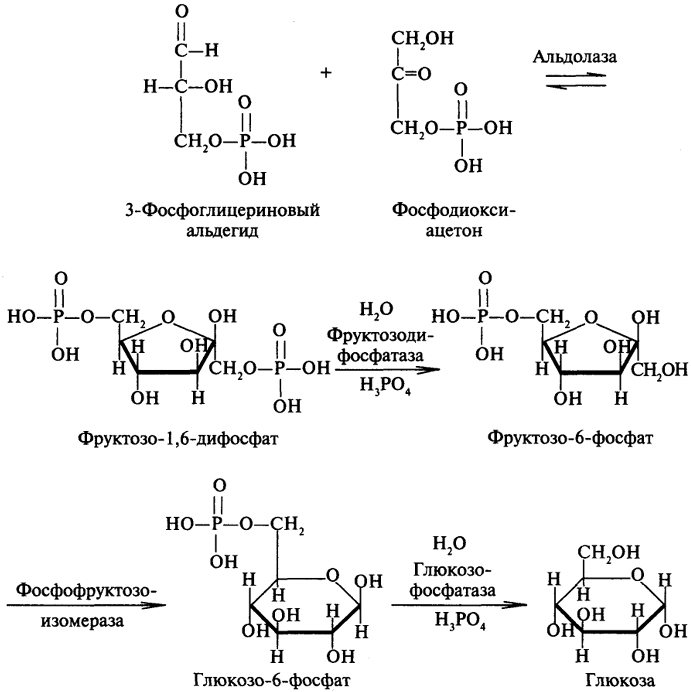
На рисунке 7-44 показаны пункты включения первичных субстратов в глюконеогенез. Большинство реакций глюконеогенеза протекает за счёт обратимых реакций гликолиза (рис. 7-45, реакции 9, 8, 7, 6, 5, 4, 2) и катализируется теми же ферментами. На этих стадиях реакции глюконеогенеза протекают другими путями.

Рассмотрим процесс синтеза глюкозы из пирувата. Образование фосфоенолпирувата из пирувата происходит в ходе двух реакций (рис. 7-45, реакции 11, 12), первая из которых протекает в митохондриях. Дальнейшие превращения оксалоацетата протекают в цитозоле. Оксалоацетат в митохондриальном матриксе восстанавливается с образованием маната (рис. 7-47) при участии NADH (обратная реакция цитратного цикла). Следует отметить, что этот обходной участок глюконеогенеза требует расхода двух молекул с макроэргическими связями (АТФ и ГТФ) в расчёте на одну молекулу исходного вещества — пирувата.
Это — пируват-карбоксилаза, фосфоенолпируваткарбоксикиназа, фруктозе-1,6-бисфосфатаза и глюкозо-6-фосфатаза. В ходе этого процесса расходуются 6 моль АТФ на синтез 1 моль глюкозы из 2 моль пирувата. Лактат, образованный в анаэробном гликолизе, не является конечным продуктом метаболизма.
Лактат, образовавшийся в интенсивно работающих мышцах или в клетках с преобладающим анаэробным способом катаболизма глюкозы, поступает в кровь, а затем в печень. В печени отношение NADH/NAD+ ниже, чем в сокращающейся мышце, поэтому лактатдегидрогеназная реакция протекает в обратном направлении, т.е. в сторону образования пирувата из лактата.
Часть пирувата, образованного из лактата, окисляется печенью до СО2 и Н2О. Энергия окисления может использоваться для синтеза АТФ, необходимого для реакций глюконеогенеза. Однако основную роль в компенсации ацидоза играют почечные механизмы с участием аммиачного буфера (см. раздел 9). Одной из причин метаболического ацидоза может быть накопление молочной кислоты.
Уровень лактата в крови — результат равновесия между процессами его образования и утилизации. У нетренированных людей лактоацидоз при физической работе возникает как следствие относительного недостатка кислорода в мышцах и развивается достаточно быстро.
При некомпенсированном лактоацидозе содержание лактата в крови увеличивается до 5 ммоль/л (в норме до 2 ммоль/л). При этом рН крови может составлять 7,25 и менее (в норме 7,36-7,44). 1 — нарушение использования пирувата в глюконеогенезе; 2 — нарушение окисления пирувата.

Кроме того, при гипоксии уменьшается синтез АТФ, что следовательно, ведёт к снижению скорости глюконеогенеза — другого пути утилизации лактата. Повышение концентрации лактата и снижение внутриклеточного рН отрицательно влияют на активность всех ферментов, в том числе и пируваткарбоксилазы, катализирующей начальную реакцию глюконеогенеза.
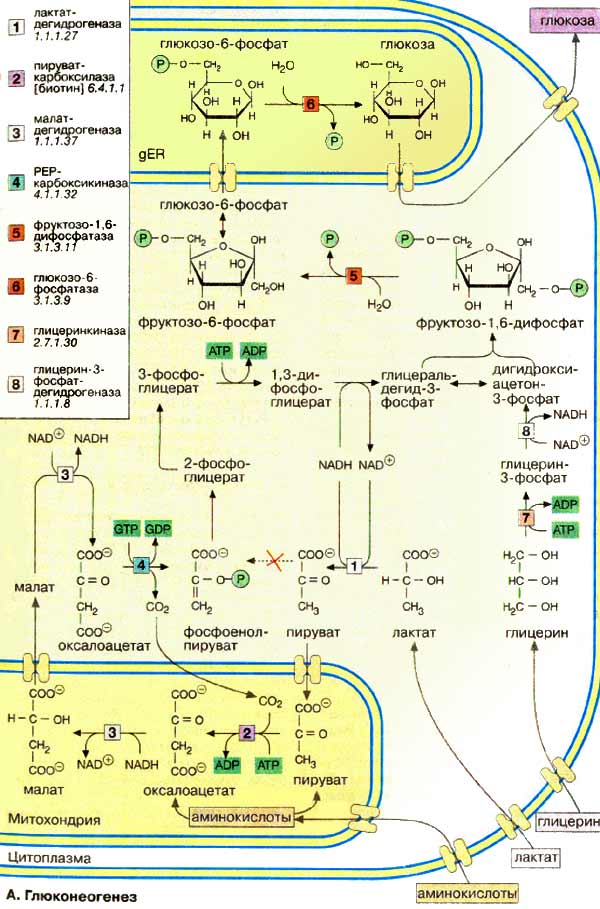
Далее пируват включается в глюконеогенез, а образовавшаяся глюкоза поступает в кровь и поглощается скелетными мышцами
В условиях голодания часть белков мышечной ткани распадается до аминокислот, которые далее включаются в процесс катаболизма. Аминокислоты, которые при катаболизме превращаются в пируват или метаболиты цитратного цикла, могут рассматриваться как потенциальные предшественники глюкозы и гликогена и носят название гликогенных.
Поэтому эти аминокислоты могут подвергнуться превращению в глюкозу и называются глюкогенными
Из всех аминокислот, поступающих в печень, примерно 30% приходится на долю аланина. Это объясняется тем, что при расщеплении мышечных белков образуются аминокислоты, многие из которых превращаются сразу в пируват или сначала в оксалоацетат, а затем в пируват. Последний превращается в аланин, приобретая аминогруппу от других аминокислот.
Отщепление фосфатной группы из фруктозо-1,6-бисфосфата и глюкозо-6-фосфата — также необратимые реакции глюконеогенеза. Рассмотрим более подробно те реакции глюконеогенеза, которые отличаются от реакций гликолиза и происходят в глюконеогенезе с использованием других ферментов.
Что еще посмотреть:
 КИШЕЧНАЯ НЕПРОХОДИМОСТЬЗадержка стула и газов - патогномоничный признак непроходимости кишечника. Эти процессы лежат в основе системной воспалительной реакции и абдоминального хирургического сепсиса, […]
КИШЕЧНАЯ НЕПРОХОДИМОСТЬЗадержка стула и газов - патогномоничный признак непроходимости кишечника. Эти процессы лежат в основе системной воспалительной реакции и абдоминального хирургического сепсиса, […] Первый способ нахождения расстояния от заданной точки до заданной прямой на плоскости.Однако чаще встречается определение расстояния от точки до прямой, в котором фигурирует длина перпендикуляра. На этом занятии мы научимся рассчитывать расстояние от точки до прямой. Это […]
Первый способ нахождения расстояния от заданной точки до заданной прямой на плоскости.Однако чаще встречается определение расстояния от точки до прямой, в котором фигурирует длина перпендикуляра. На этом занятии мы научимся рассчитывать расстояние от точки до прямой. Это […] Тема, задача, материалОлимпиады является наиболее распространенной и массовой формой организации научно-исследовательской работы студентов в университете.
Если же студент вспоминает про НИР в конце семестра […]
Тема, задача, материалОлимпиады является наиболее распространенной и массовой формой организации научно-исследовательской работы студентов в университете.
Если же студент вспоминает про НИР в конце семестра […]