 Две дисульфидные связи образованы между остатками цистеина в положениях 183-189 и 53-165. По механизму их синтеза и биологическим функциям эти гормоны объединяют в 3 группы. Регуляция синтеза и секреции гормона роста осуществляется множеством факторов. В передней доле гипофиза синтезируются гормоны, которые по химическому строению являются пептидами и гликопротеинами. Гипофиз секретирует большое количество гормонов, участвующих в регуляции различных биохимических процессов и физиологических функций.
Две дисульфидные связи образованы между остатками цистеина в положениях 183-189 и 53-165. По механизму их синтеза и биологическим функциям эти гормоны объединяют в 3 группы. Регуляция синтеза и секреции гормона роста осуществляется множеством факторов. В передней доле гипофиза синтезируются гормоны, которые по химическому строению являются пептидами и гликопротеинами. Гипофиз секретирует большое количество гормонов, участвующих в регуляции различных биохимических процессов и физиологических функций.
Многие эндокринные железы вырабатывают несколько гормонов, имеющих различное строение и осуществляющих различные функции. Гипоталамус занимает важнейшее место в иерархической системе, объединяя высшие отделы ЦНС и эндокринные железы.
В группу гормонов, относящихся к гликопротеинам, входят также гонадотропные гормоны гипофиза ЛГ и ФСГ и хорионический гонадотропин (ХГ) (рис. 11-14)
Тиреолиберин разрушается в клетках-мишенях и в крови под действием специфических протеаз. Рис. 11-9. Структура некоторых гормонов гипоталамуса. В передней доле гипофиза кортиколиберин увеличивает синтез и секрецию проопиомеланокортина и образование кортикотропина. Биологической активностью обладает и ациклическая восстановленная форма пептида. В тканях соматостатин присутствует в форме пептида, содержащего 28 аминокислотных остатков и может служить предшественником пептида, состоящего из 14 аминокислотных остатков.
Подобно другим пептидным гормонам, соматостатин взаимодействует с рецепторами плазматической мембраны клеток. Все типы рецепторов экспрессируются в передней доле гипофиза и гипоталамусе и обладают различной степенью сродства к разным структурным формам соматостатина. Рецепторы к соматостатину присутствуют во многих опухолевых клетках, секретирующих гормоны.
Секреция гормонов гипофиза обусловлена сочетанием нервных и гуморальных сигналов. При этом один и тот же агонист (например, норад-реналин) может вызывать противоположные изменения в секреции гипофизарных гормонов. С другой стороны, секреция каждого гормона может контролироваться многочисленными факторами. Гормон роста синтезируется в соматотроф-ных клетках, наиболее многочисленных в передней доле гипофиза.
В крови гормоны не связаны с нейрофизином
Уровень гормона роста в плазме крови не превышает 3 нг/мл. Секреция гормона роста носит пульсирующий характер с интервалами в 20-30 мин. Один из самых больших пиков отмечается вскоре после засыпания.
Участвует в регуляции энергетического и минерального обмена. Рис. 11-10. Гормон роста человека. Первичные эффекты гормона роста кратков-ременны и инсулиноподобны. Основное действие гормона роста направлено на регуляцию обмена белков и процессов, связанных с ростом и развитием организма.
Инсулиноподобные факторы роста оказывают своё действие различными путями: эндокринным, паракринным и аутокринным (рис. 11-13). Это указывает на то, что инсулиноподобные факторы роста более активны в отношении их действия на рост и развитие клеток. Под влиянием гормона роста увеличивается ширина и толщина костей, и одновременно с этим ускоряется рост других тканей, включая соединительную ткань, мыпщы и внутренние органы.

Он обладает более высоким сродством к Т3 и Т4 и в нормальных условиях связывает почти всё количество этих гормонов
Пролактин близок по химическому строению гормону роста. Он состоит из 199 аминокислотных остатков, образующих одну полипептидную цепь с тремя дисульфидными связями. Рецепторы пролактина присутствуют в клетках многих тканей: в печени, почках, надпочечниках, яичках, яичниках, матке и других тканях.

Все 3 гормона имеют общие антигенные детерминанты и обладают рост-стимулирующей и лакто-генной активностью. Существует гипотеза, согласно которой гены этих гормонов возникли в результате дупликации одного гена-предшественника. Стимуляция секреции тиреотропина происходит под влиянием тиреолиберина, а основное ингибирующее действие оказывает повышение уровня тиреоидных гормонов. Трансдукция сигнала тиреотропина в клетки щитовидной железы происходит через рецепторы плазматической мембраны и активацию аденилатциклазы.
В гипофизе человека найдены β-липотропин, γ-липотропин и β-эндорфин
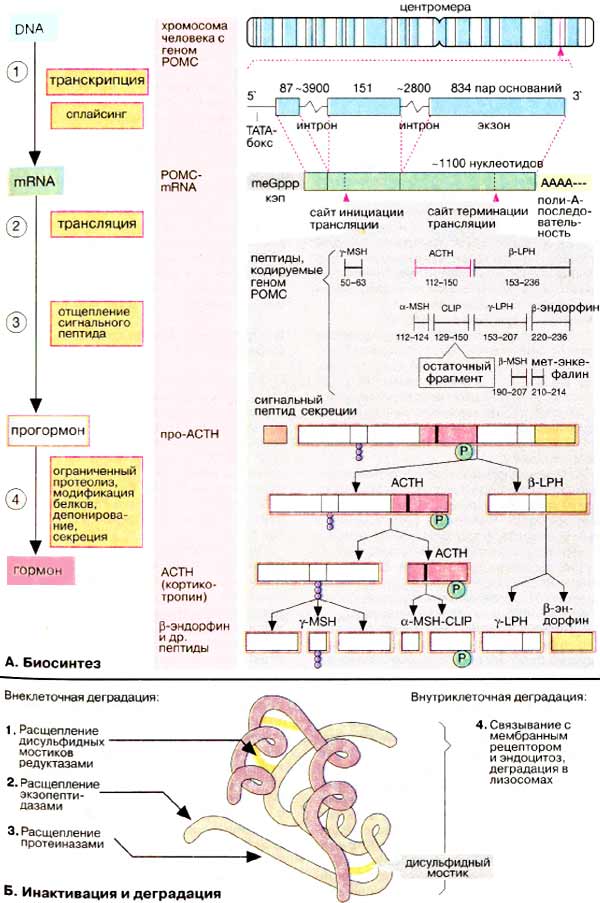
Некоторые иммуноглобулины класса G, взаимодействуя с рецепторами тиреотропина, имитируют эффекты гормона. После отщепления сигнального пептида происходит частичный протеолиз оставшейся полипептидной цепи с образованием АКТГ и β-липотропина (β-ЛП).
МСГ и кортикотропиноподобный гормон промежуточной доли у человека практически не образуются, так как у взрослых людей промежуточная доля не развита. V2-рецепторы расположены в клетках почечных канальцев. Рис. 11-16. Структура вазопрессина и окситоцина.

У большинства животных и человека в положении 8 вазопрессина находится аргинин вместо лизина, в связи с чем он обозначается как аргинин вазопрессин. В некоторых случаях гипосекреция гормона роста может быть результатом аутоиммунного повреждения соматотрофных клеток гипофиза, черепно-мозговой травмы или радиации. Гиперфункция гормона роста обычно возникает в,результате образования гормонпродуцирующей опухоли соматотрофных клеток гипофиза, что приводит к повышению ростовой активности.
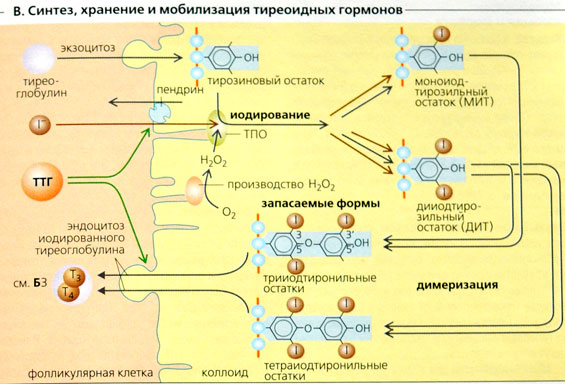
Вследствие этого развиваются продолжительная активация аденилатциклазы, избыточное образование цАМФ и избыточная секреция соматотропного гормона. В щитовидной железе синтезируются гормоны — йодированные производные тирозина. К ним относят 3,5,3′-трийодтиронин (трийодтиронин, Т3) и 3,5,3′,5′-тетрайодтиронин (Т4), или тироксин (рис. 11-17).
Кроме того, секреция гормонов гипоталамуса и гипофиза регулируется по механизму обратной связи гормонами, продукцию которых они стимулируют в органах-мишенях. Основная биологическая функция тиреотропина — стимуляция синтеза и секреции йод-тиронинов (Т3 и Т4) в щитовидной железе.
Что еще посмотреть:
 Правила пожарной безопасности на складах ГСМ2.2.13. При работах по перекачке топлива должны соблюдаться правила техники безопасности и пожарной безопасности в соответствии с разделом 13.4 Руководства. Осмотр трассы можно производить […]
Правила пожарной безопасности на складах ГСМ2.2.13. При работах по перекачке топлива должны соблюдаться правила техники безопасности и пожарной безопасности в соответствии с разделом 13.4 Руководства. Осмотр трассы можно производить […] Под световым микроскопом казалось, что клетка заполнена чем-то вроде жидкой плазмы или золя, в котором «плавают» ядро и другие органоидыПод световым микроскопом казалось, что клетка заполнена чем-то вроде жидкой плазмы или золя, в котором «плавают» ядро и другие органоиды. К элементам цитоскелета относят белковые […]
Под световым микроскопом казалось, что клетка заполнена чем-то вроде жидкой плазмы или золя, в котором «плавают» ядро и другие органоидыПод световым микроскопом казалось, что клетка заполнена чем-то вроде жидкой плазмы или золя, в котором «плавают» ядро и другие органоиды. К элементам цитоскелета относят белковые […] На эмоционально-экспрессивную окраску слова влияет его значениеНа эмоционально-экспрессивную окраску слова влияет его значение. Нейтральные в эмоционально-экспрессивном отношении слова обычно относятся к общеупотребительной лексике (хотя это и не […]
На эмоционально-экспрессивную окраску слова влияет его значениеНа эмоционально-экспрессивную окраску слова влияет его значение. Нейтральные в эмоционально-экспрессивном отношении слова обычно относятся к общеупотребительной лексике (хотя это и не […]